This horse is in the process of losing its pigment.
It will end up being all white by the time it is eight years old.
Image: Horse Wallpaper [larger view].

Even though I have always been a fan of black horses, my heart did leap at the sight of the noble Shadowfax racing towards Gandalf in response to his call in the Lord of the Rings. White horses have symbolized purity throughout most ages and cultures, so political leaders have often been portrayed astride a white horse. Interestingly, these so-called white horses are actually Gray. White horses have pink skin and blue eyes. Gray horses are typically born black, bay or chestnut in color but rapidly lose all hair pigmentation as they age, ending up completely white by the time they are eight years old. Meanwhile, their skin remains black and their eyes are typically dark, giving a silvery or grey cast to their seeming snowy-whiteness. This popular coat color results from an autosomal dominant mutation to a gene that was just identified by a research team at Uppsala University in Sweden. Unfortunately, this coat color is also linked to an increased risk of developing a specific form of cancer: melanoma. Thus, this finding has important implications for human medical research.
"It is a fascinating thought that once upon a time a horse was born that turned Gray and subsequently white and the people that observed it were so fascinated by its spectacular appearance that they used the horse for breeding so that the mutation could be transmitted from generation to generation," reports geneticist Leif Andersson of the University of Uppsala in Sweden who led the large international research team.
Based on genetics, it has been previously speculated that rapid graying in horses was due to an autosomal dominant gene, but the identity of that gene has remained elusive. Previous research suggested that this gene might be located on a specific region of chromosome 25.
To identify this gene, the researchers analyzed the DNA from 727 Gray horses and 131 nongrays. The region of the Gray-causing mutation contains four genes:
nuclear receptor subfamily 4, group A, member 3 (NR4A3), syntaxin-17 (STX17), thioredoxin domain-containing-4' (TXNDC4) and inversin (INVS) -- none of which had previously ever been associated with pigment defects or melanoma. Even more interesting, they found that all of the Gray horses had a 4600 base pair duplication in intron 6 (a noncoding region) of one particular gene, STX17, which was not present in nongray horses (figure 2);
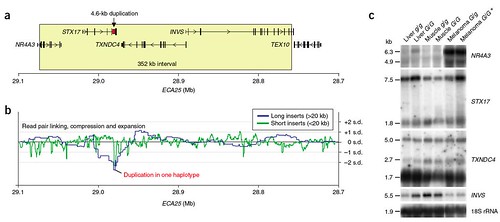
Figure 2: Molecular characterization of the locus of the Gray-causing mutation in horses. (a) Gene content of the mutation interval. The 352-kb region showing complete association with the Gray phenotype is indicated by a box; the location of the 4.6-kb duplication in STX17 intron 6 is marked with an arrow. The annotation is based on the horse genome assembly as presented on the UCSC server (built Jan. 2007, EquCab1 assembly). (b) Average paired-end read compression and expansion in the horse genome assembly across the region in standard deviations (s.d.), broken down by insert size. Green, 4.5-10 kb (plasmids); blue, 40 kb (fosmids) and 180 kb (BAC ends). The only spot in which both short and long inserts are significantly compressed (by more than two s.d.) coincides with the 4.6-kb duplication in STX17 intron 6. (c) Multiple-tissue northern blot analysis of genes in the mutation interval; 18S rRNA was used as an internal control. G, Gray mutant allele; g, wild-type allele. *, mRNA from a melanoma cell line derived from a heterozygous (G/g) horse (M.H.S., unpublished data) is shown in this lane for hybridization with the NR4A3 probe. The estimated transcript sizes are given to the left [larger view].
DOI: 10.1038/ng.185.
According to the data, this duplication (figures 2a and 2b) does not affect the size of the final gene product, but it is associated with overproduction of STX17 and of the nearby gene, NR4A3, especially in the case where melanoma has developed (figure 2c).
Because this duplication was found only in Gray horses, the team concluded that it is the cause of the Gray color, even though STX17 has no known function. When the team screened Gray and nongray horses from 14 breeds [Arabian, Connemara, Fjord, Friesian, Haflinger, Icelandic, Lipizzaner, Morgan, New Forst pony, North Swedish, Sheltand pony, Swedish Warmblood, Thoroughbred, Welsh], they found this identical mutation in all Gray horses, indicating that they all had a common ancestor (data not shown). For example, all Gray thoroughbreds have one Gray heterozygous mare, Twilight, in their ancestry. In fact, careful scrutiny of the pedigrees of these horse breeds reveals that Arabian horses are somewhere in all of their ancestries, indicating that the original mutation occurred in an Arabian horse. Considering our fascination with animals that lack pigment (look at all our common barnyard animals for numerous examples), it is not difficult to conclude that ancient humans selectively bred horses for this unique color.
After determining that the Gray mutation is fully dominant, the researchers report that there are several characteristics that differ between Gray individuals: the speed of graying, amount of pigmented speckling present (commonly known as "flea-bitten grey", see C, below), incidence of melanomas and the development of a vitiligo-like skin depigmentation -- all of which show considerable variation among Gray horses. To determine if these characters are related to the number of copies of the Gray gene, the researchers then measured these variations in 694 Gray Lipizzaner horses (467 G/g and 227 G/G) (figure 3);
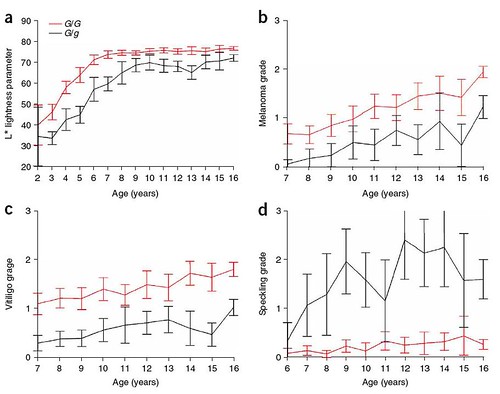
Figure 3: Analysis of phenotypic differences between heterozygous (G/g; black lines) and homozygous (G/G; red lines) Gray Lipizzaner horses. Shown are degree of lightness (a), measured as light reflectance, and grades of melanoma (b), vitiligo (c) and speckling (d). The analysis included 467 G/g and 227 G/G Lipizzaner horses, which were genotyped for the STX17 duplication by a PCR-based method. The data show means ± 2 s.e.; that is, nonoverlapping bars indicate a statistically significant difference. Overall P < target="window" href="http://farm4.static.flickr.com/3082/2734493084_321df34e88_o.jpg" width="911" height="718">larger view].
DOI: 10.1038/ng.185.
The researchers found that homozygotes (G/G) showed a faster rate of graying and ended up with more consistent depigmentation (figure 3a), had a higher chance of developing vitiligo (figure 3c), and ended up with little or no speckling (figure 3d; also refer to C, below). Worst, homozygotes also had a much higher incidence of melanoma (figure 3b).
Interestingly, some horses develop large pigmented areas, known as "blood marks" (see D, below). These are heterozygotes where the duplicated region has been deactivated in some regions of their bodies so normal pigment production has been able to resume.

(c) Gray horse with characteristic speckling (numerous small spots of pigmented hair, also called 'flea-bitten' Gray). Photo used with permission of Monika H. Seltenhammer. (d) Gray horse with red 'blood marks'. There is notable contrast between the remaining black pigmentation in the areas showing graying and the red pigmentation in the blood marks. Photo used with permission of Emilie Kajle. [larger view].
DOI: 10.1038/ng.185.
Since roughly 75% of Gray horses that are older than 15 years of age develop a benign form of melanoma that can transform into a malignancy, the Gray horse presents medical researchers with an interesting model for learning more about the molecular pathway that leads to tumor development in humans. So the team identified precisely what this Gray mutation looks like and investigated how it upregulates the production of the two neighboring genes, STX17 and NR4A3. They also investigated whether this mutation is present in three human melanoma cell lines (figure 4);
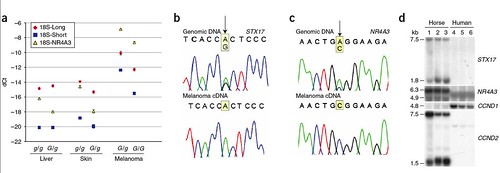
Figure 4: Expression analysis of STX17 and NR4A3. (a) Real-time PCR analysis showing expression of STX17 (short and long isoforms) and NR4A3 in relation to the expression of 18S rRNA in various tissues from Gray and non-Gray horses. dCt, difference in Ct value (threshold cycle) for the control gene (18S in this case) and the target transcript. (b,c) Differential expression analysis for STX17 (b) and NR4A3 (c) using melanoma tissue from G/g heterozygotes; the nucleotide sites for the SNPs in STX17 and NR4A3 correspond to positions 28,972,811 bp (intron 6, 5' UTR of the alternative transcript) and 29,063,351 bp (exon 8), respectively. Genomic DNA was used as a reference. (d) Northern blot analysis showing that enhanced expression of STX17 and NR4A3 are associated with high expression of CCND2 but not CCND1 in Gray melanomas. 1, melanoma tissue from G/g horse; 2, melanoma cell line from G/g Lipizzaner horse; 3, melanoma cell line from G/g Arabian horse; 4, human melanoma cell line A375; 5, human melanoma cell line M5; 6, human melanoma cell line BL. The horse melanoma cell lines were established by M.H.S. (unpublished data) [larger view].
DOI: 10.1038/ng.185.
Since none of the resulting gene products has been altered in size, the protein structures are not obviously altered. However, northern blot analysis confirms that the amount of the two genes produced is dramatically increased (figure 4a; also see figure 2c). This was identified as a cis-acting regulatory mutation that underlies the upregulation of expression (fugure 4c). A cis mutation acts on the gene in which it is located, or upon nearby genes, and a regulatory gene mutation either allows a gene to be produced when it normally would not be, or to be produced in greater quantities than normal.
"Since the first description of melanomas in grey horses in 1903, researchers have questioned how a mutation causing loss of hair pigmentation can also cause melanomas. Our results suggest a possible explanation," reports Andersson.
Because human melanomas are characterized by an overproduction of the cyclin D1 (CCND1) and cyclin D2 (CCND2) gene products, the team investigated whether these genes' expression profiles were affected similarly in Gray horses. They found that Gray horse melanoma tissue and cell lines both showed high expression of CCND2 but not CCND1 (figure 4d) but this was different from three human melanoma cell lines. The three human melanoma cell lines that were investigated had low expression of STX17, NR4A3 and CCND2 but high expression of CCND1.
Currently, the team does not know if one, or both STX17 and NR4A3 are responsible for the loss of hair pigmentation and melanoma growth, but it is possible that the Gray mutation has other effects.
"We propose that the Grey mutation stimulates growth of melanocytes and that this leads to a premature loss of the melanocyte stem cells needed for hair pigmentation whereas the mutation promotes an expansion of some of the melanocytes causing skin pigmentation," Andersson speculates.
Even though the Gray mutation doesn't affect the structure of the proteins themselves, it provides a beautiful example of the regulatory power of mutations as a major underlying mechanism for phenotypic diversity within and between species. These results show that only one copy of the Gray mutation is necessary to exert profound molecular effects upon the regulation of two neighboring genes.
"It is very likely that regulatory mutations like the one we found in these white horses constitute the dominating class of mutations explaining differences between breeds of domestic animals as well as between species like humans and chimpanzee," speculates Andersson.
Source
Rosengren Pielberg, G., Golovko, A., Sundström, E., Curik, I., Lennartsson, J., Seltenhammer, M.H., Druml, T., Binns, M., Fitzsimmons, C., Lindgren, G., Sandberg, K., Baumung, R., Vetterlein, M., Strömberg, S., Grabherr, M., Wade, C., Lindblad-Toh, K., Pontén, F., Heldin, C., Sölkner, J., Andersson, L. (2008). A cis-acting regulatory mutation causes premature hair graying and susceptibility to melanoma in the horse. Nature Genetics, 40(8),
No hay comentarios:
Publicar un comentario